
Классификация ядерных рецепторов и их лиганды
Функциональный синергизм
Коактиваторы этого семейства взаимодействуют преимущественно с активационной функцией 2 (AF-2) Рц, при этом обеспечивается функциональный синергизм AF-2 и AF-1. Кроме того, эти белки способны взаимодействовать с основными транскрипционными факторами (например, TFIIB и ТВР). Члены этого семейства (SRC-1,2,3) кодируются 3 генами, причём каждый из членов представлен несколькими сплайсинговыми вариантами. Структурные различия наблюдаются преимущественно в С-концевой области и касаются, в частности, размера полиглутаминовых фрагментов. Консенсусные мотивы LXXLL (L — лейцин, X — любая аминокислота) за счёт формирования гидрофобной поверхности из остатков лейцина на амфипатической сс-спирали обеспечивают связывание коактиватора с гидрофобной бороздкой Рц, формируемой в присутствии агониста несколькими фрагментами молекулы при обязательном участии 12-й а-спирали. В присутствии антагониста эта а-спираль, напротив, становится стерическим препятствием для взаимодействия Рц с LXXLL мотивом коактиватора. Наличие нескольких таких мотивов в одной молекуле коактиватора даёт возможность этой молекуле контактировать одновременно с обеими молекулами Рц в составе димеров. Аналогичные мотивы имеются и во многих других корегуляторах ядерных Рц. Вариации в этих мотивах могут определять избирательность связывания корегуляторов с определёнными Рц.
К коактиваторам ядерных Рц (кроме членов семейства SRC) относятся и некоторые другие белки, в частности ЕЗ убиквитин-про-теинлигазы Е6-АР (см. ниже) и RPF-1 (retina-derived POU-domain factor-1 — содержащий домен POU фактор 1 сетчатки). Коактиватор SUG-1 не содержит LXXLL-мотивов. По-видимому, SUG-1 взаимодействует с фрагментами Рц, отличными от AF-2 (см. ниже). Коактиватор L7/SPA (SPA — switch protein for antagonists — белок, переключающий на антагонисты) усиливает активность частичных агонистов гормонов, но не влияет на активность полных агонистов и полных антагонистов.
Многокомпонентный коактиваторный белковый комплекс
Не исключено, что этот (или сходные) коактиватор связан с действием селективных аналогов гормонов. Многокомпонентный коактиваторный белковый комплекс TRAPs/ DRIPs (thyroid hormone receptor associated proteins — белки, ассоциированные с Рц тиреоидных гормонов/vitamin D3 receptor-interacting proteins — белки, взаимодействующие с Рц витамина D3) взаимодействует с Рц через TRAP220/PBP (РВР — PPAR binding protein — белок, связывающий PPAR). В реализацию эффектов ядерных Рц могут быть вовлечены и другие факторы широкого спектра действия, в частности позитивные кофакторы (PCs). PGC-1 (коактиватор-1 PPARy), в отличие от большинства других коактиваторов, взаимодействует не только с AF-2, но и с областью, локализованной в шарнирном домене Рц. Этот коактиватор, как и сам PPARy, преобладает в буром жире и скелетных мышцах и зависит от температурного режима, что может указывать на ключевую роль этого коактиватора в термогенезе. Экспрессию PGC-1 в печени (необходима для индукции глюконеогенеза) индуцируют голодание и недостаточность инсулина. Кроме того, этот коактиватор способен взаимодействовать с факторами сплайсинга. В избирательность действия ядерных Рц может вносить вклад их селективное связывание с группой белков (TAFus — TBP-associated factor), ассоциированных с белком ТВР, связывающим ТАТА-последовательность в промоторе контролируемого гена. Полагают, что благодаря избирательности взаимодействия с разными TAFns разные ядерные Рц могут способствовать формированию различающихся вариантов комплекса транскрипционного фактора TFIID.
Более селективные коактиваторы включают SRA (steroid receptor RNA activator — РНК-активатор Рц стероидов). Этот небелковый (РНК) коактиватор избирательно усиливает активность AF-1 Рц стероидов. По-видимому, SRA служит мостиком между AF-1 Рц и ассоциированными с AF-2 коактиваторами (SRC) посредством взаимодействия с коактиваторными белками р72/р68, содержащими участки связывания SRA и SRC.
Белковые комплексы
Представители группы коактиваторов ARA (от androgen receptor-associated protein — ассоциированный с Рц андрогенов белок) избирательно взаимодействуют с AF-1 или AF-2 Рц андрогенов и — что особенно интересно — способны модулировать агонистическую активность частичных агонистов/ антагонистов, т.е. действуют подобно коактиватору L7/SPA (см. выше). Белковые комплексы группы TIF-1/SL1 (transcriptional intermediary factor 1 — транскрипционный промежуточный фактор 1) могут опосредовать действие ядерных Рц на промоторы рДНК, причём опосредующий эффект зависит от ацетилирования TIF-1. Особую нишу среди коактиваторов занимают белки высокомобильной группы (high mobility group protein) HMG-1 и HMG-2, которые избирательно повышают ДНК-связывающую активность Рц стероидных гормонов за счёт образования стабильных тройных комплексов Рц-HMG—ДНК. Предполагают, что HMG функционально заменяет необходимое для взаимодействия Рц с малой бороздкой ДНК С-концевое расширение ДНК-связывающих доменов других ядерных Рц, которое отсутствует в Рц стероидов.
Коинтеграторы СВР и рЗОО. СВР (белок, связывающий CREB — белок, связывающий цАМФ-чувствительный элемент) и рЗОО служат коактиваторами не только ядерных Рц, но и ряда других активаторов транскрипции. СВР может одновременно взаимодействовать с ядерными Рц и рядом коактиваторов, включая члены семейства SRC, а также с гистонацетилазами. В соответствии с гипотезой «выстрела и перезарядки» рЗОО и Рц кооперативно повышают транскрипцию лишь на первом раунде, а для повторной сборки транскрипционного комплекса необходим лишь Рц. Есть основания полагать, что in vivo ряд коактиваторов находится в виде преформированных относительно стабильных олигомерных комплексов, которые рекрутируются активированным Рц как единое целое и в свою очередь могут рекрутировать дополнительные корегуляторы, образуя с ними более рыхлую связь.
Образование комплексов Рц
Таким образом, связывание агониста стимулирует образование комплексов Рц с одним или более коактиваторами, такими как коактиватор Рц стероидов (SRA); фактор, ассоциированный с рЗОО/СВР (p/CAF), СРВ/рЗОО. Коактиваторы SRC-1, p/CAF, СВР обладают собственной гистонацетилазной активностью. Кроме того, эти и другие коактиваторы рекрутируют гистонацетилазы с иной субстратной специфичностью. В обоих случаях ацетилирование гистонов ведёт к релаксации и соответственно дерепрессии хроматина за счёт ослабления взаимодействия гистонов с отрицательно заряженными группами ДНК и ослабления связей между нуклеосомами. Кроме гистонов, субстратами для регулируемых ядерными Рц ацетилтрансфераз могут служить транскрипционные факторы (например, белок р53), модификация которых ведёт к изменению их активности. Сходным (хотя и с противоположным конечным результатом) образом могут действовать обладающие АТФазной активностью белковые комплексы NURD (complex containing nucleosome remodeling and histone deacetylase activities — комплекс, содержащий ремоделируюшую нуклеосомы и гистондеацетилазную активность) и Mi-2. Предполагается, что эти белки нарушают структуру хроматина для обеспечения последующего доступа корепрессорам, ассоциированным с гистондеацетилазами, которые, напротив, фиксируют неактивную форму хроматина. Белок NSD-1 (nuclear receptor-binding SET domain-containing protein), взаимодействующий с ядерными Рц и содержащий домен SET, содержит как активационные, так и репрессорные домены и, по-видимому, может стабилизировать или релаксировать структуру хроматина в зависимости от стадии развития.
Нуклеосома может не только препятствовать транскрипционной активности гена, но и при соответствующем позиционировании повышать эту активность.
Оборачивание ДНК вокруг гистонового ядра
Оборачивание ДНК вокруг гистонового ядра может приводить к пространственному сближению дистальных регуляторных участков гена (в том числе и гормончувствительных элементов) с проксимальными участками, взаимодействующими с базовыми факторами транскрипции. В результате ассоциированные с гормончувствительным элементом ядерные Рц получают возможность прямо или опосредованно взаимодействовать с базовыми транскрипционными факторами и тем самым стабилизировать преинициативный транскрипционный комплекс.
Позиционирование нуклеосом служит одним из способов придания действию гормонов тканевой специфичности. Например, эстрогенчувствительный элемент гена пролактина локализован вне нуклеосомы только в клетках, секретирующих этот гормон (эстрогенчувствительный элемент в нуклеосоме недоступен для Рц эстрогенов).
Корепрессоры. Ядерные Рц типа II и некоторые другие способны снижать базальную активность контролируемых промоторов генов в отсутствие гормонального лиганда. Эта способность определяется взаимодействием Рц с корепрессорными белками. Структурно-функциональная организация корепрессоров представлена на примере NcoR (корепрессора ядерных Рц). Сходную организацию при значительной гомологии последовательности имеет ко-репрессор SMRT (silencing mediator of retinoic acid and thyroid hormone receptor — посредник молчания Рц ретиноидных и тиреоидных гормонов), за исключением того, что он не содержит последовательностей, соответствующих репрессорным доменам RDI и RDII.
Субдомены А и В домена SANT (области гомологии белков SWI3, ADA2, N-CoR и TFIIIB) играют важную роль в поддержании непермиссивной для транскрипции среды путём деацетилирования гистонов. Субдомен А связывает и активирует гистондеацетилазу 3 (HDAC3), а субдомен В участвует в связывании корепрессора с гистоном и направлении HDAC к субстрату. Кроме того, этот субдомен ингибирует гистонацетилтрансферазы.
Связывание тримерного Рц
В отсутствие лиганда корепрессор NCoR (благодаря двум доменам взаимодействия с ядерными Рц [ID-N и ID-С], расположенным на С-конце NCoR и включающим а-спиральные элементы с мотивом LXXI/HIXXXI/L) может одновременно взаимодействовать с двумя рецепторными молекулами, формирующими димер на гормончувствительном элементе.
Рядом с ID-N был идентифицирован ещё один ID с центральной последовательностью IDVII. Замена остатков лейцина в IDs коактиваторов на изолейцин в IDs корепрессоров обеспечивает предпочтительное связывание последних с апорецепторами, а не с Рц, находящимися в комплексе с лигандами. Остаётся неясным, зачем нужен дополнительный сайт связывания Рц на молекуле корепрессора NCoR. Возможно, его необходимость определяется известной избирательностью отдельных IDs в отношении разных Рц. Не исключено также, что дополнительный ID может обеспечивать связывание тримерного Рц, который, как предполагается, может формироваться на некоторых негативных HREs (см. ниже).
Области взаимодействия с корепрессором локализованы на N-и С-концах лигандсвязывающего домена Рц (в некоторых Рц также, возможно, в шарнирном домене) и представляют собой гидрофобную ложбинку, формируемую несколькими а-спиралями (НЗ-Н4 и НИ—HI2), которая частично перекрывает поверхность взаимодействия Рц с коактиваторами. В случае регулируемых лигандами Рц 12-я а-спираль играет особую роль: изменение её пространственной ориентации при связывании лиганда становится стерическим препятствием для взаимодействия Рц с корепрессором, и
Рц приобретает способность взаимодействовать с коактиватором.
Н12 в RXR (партнёре димеризации ряда других ядерных Рц) выполняет, по-видимому, ещё одну функцию: в ano-RXR эта а-спи-раль служит стерическим препятствием для связывания корепрессора, а в гетеродимерах (TR/RXR) в отсутствие лигандов ориентация Н12 за счёт участия в межрецепторных взаимодействиях меняется таким образом, что это ведёт к связыванию корепрессора с димером.
- Предыдущие статьи из рубрики: Эндокринология
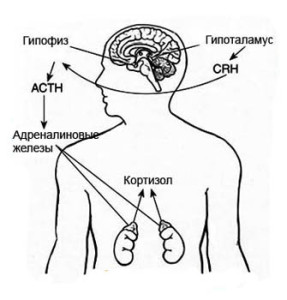 Общие свойства рецепторов
Общие свойства рецепторовРецепция и проведение сигнала Эти системы могут включать так называемые «вторые посредники» — низкомолекулярные соединения (циклические нуклеотиды, фосфолипиды […]
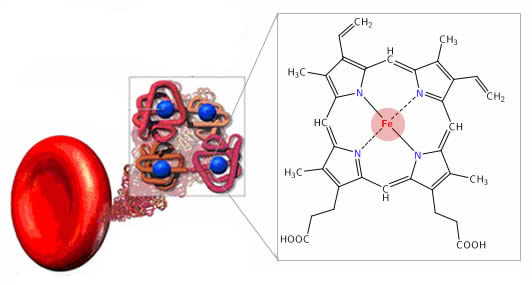 Функции транспортных белков
Функции транспортных белковФункции белков Функции белков, связывающих ИФР, связанные с ИФР торможение взаимодействия ИФР с Рц за счёт конкуренции за […]
 Транспорт сигнальных соединений
Транспорт сигнальных соединенийТранспортные белки и их лиганды К известным транспортным белкам относятся белки, связывающие стероиды (1), тиреоидные гормоны и ретиноиды […]
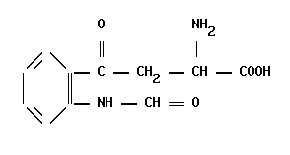 Производные триптофана
Производные триптофанаЗавершение процессов биосинтеза Так как конвертазы экспрессируются тканеспецифично, то и набор продуктов процессинга ПОМК в передней доле гипофиза, […]
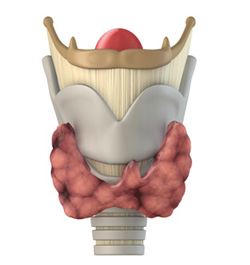 Тиреоидные гормоны
Тиреоидные гормоныЙодирование тироглобулина происходит в комплексе Гольджи у апикальной поверхности клеток под действием тиропероксидазы — гемсодержащего белка, катализирующего 1) […]