
Рецепторы, сопряжённые с g-белками
Терминация рецепторного цикла
Выявлено взаимодействие GPy с рядом других ионных каналов. К таким каналам относятся Са-каналы L-типа, потенциалзависимый К+-канал Kvl.l, К+-канал IRK3, АТФ-чувствительные каналы (Кдтр). КАТР выполняют защитную функцию при метаболическом стрессе, регулируют секрецию инсулина и возбудимость сердца. Связывание GPy с СЕ SUR этих каналов снимает тормозное действие АТФ на проводимость КАТр.
Активация Рц в результате связывания агониста ведёт не только к обмену ГДФ на ГТФ в G-белке, диссоциации а- и Ру-СЕ G-белка и их действию на эффекторы, но и к включению процессов, ведущих к десенситизации Рц. Один из путей десенситизации — фосфорилирование Рц-зависимой от второго посредника киназой (например, цАМФ-зависимой протеинкиназой или протеинкиназой С). Эта форма регуляции не требует присутствия агониста, а зависит от активности киназ. Полагают, что такое фосфорилирование вызывает изменение конформации рецепторной молекулы, в результате чего сродство Рц к G-белку снижается. Второй тип киназ, фосфорилирующих Рц — GRKs (киназы Рц, сопряжённых с G-белками). Эти киназы участвуют в гомологичной десенситизации Рц, поскольку только в присутствии агониста данный Рц служит субстратом для GRK. К настоящему времени клонировано 7 GRKs. GRK1 — киназа родопсина. GRK2 и GRK3 экспрессируются во многих тканях и фосфорилируют многие Рц. Нокаут GRK2 у мыши приводит к гибели эмбрионов, а у гетерозигот обнаруживаются нарушения сердечной сократимости. У мышей с нокаутированным геном GRK3 нарушена десенситизация обонятельных Рц. Нокаут GRK5 ведёт к гиперчувствительности мускариновых Рц (включая такие проявления, как слюноотделение и тремор). GRK4 экспрессируется преимущественно в семенниках, GRK6 — во многих тканях, a GRK7 — только в структурах глаза.
Аналог зрительного аррестина
Помимо фосфорилирования, для десенситизации Рц требуется по меньшей мере ещё один белок — аналог зрительного аррестина, участвующего в десенситизации родопсина. Клонировано 3 таких белка — р-аррестины 1 и 2, экспрессируемые повсеместно, и аррестин колбочек, локализация которого ограничена колбочками сетчатки и эпифизом. Наибольшая концентрация р-аррестинов найдена в постсинаптических структурах. Р-Аррестины действуют в качестве адаптеров, связывающих фосфорилированные Рц с покрытыми клатрином окаймлёнными ямками, подвергающимися далее эндоцитозу. Помимо клатрина, р-аррестины взаимодействуют со вторым структурным элементом окаймлённых ямок — гетеротетрамерным комплексом АР2, СЕ которого называются адаптинами. р-Аррестины взаимодействуют также с фосфолипидами мембран. Вскоре Рц (в комплексе с аррестином или без него) ин-тернализуются в виде эндосом и либо подвергаются дефосфорили-рованию и возвращению в плазматическую мембрану, либо деградации. Существуют и другие пути интернализации Рц, не зависящие от клатрина и р-аррестина (например, через кавеолы). Рц разных семейств могут различаться по соотношению сродства к двум р-аррестинам. Кроме того, в случае одних Рц (например, Р2-АдРц) (группа 1) после эндоцитоза р-аррестин быстро диссоциирует от Рц, а в других случаях (например, Рц АТ1А ангиотензина) (группа 2) остаётся в комплексе с Рц и после эндоцитоза. Данные особенности Рц определяются структурой их С-концевой последовательности. Прочность связывания р-аррестина с Рц обратно коррелирует со скоростью рециклизации Рц, т.е. дефосфори-лирования и возвращения в плазматическую мембрану.
Вызываемая р-аррестинами десенситизация Рц и стимулируемая Р-аррестинами интернализация — независимые процессы. Например, мускариновый Рц М2 ацетилхолина десенситизируется с участием р-аррестина, но интернализуется с помощью механизма, независимого от р-аррестина. Интернализация Рц может служить как формой временной десенситизации (за счёт блокирования доступа сигнального соединения к Рц), так и этапом в ресенситизации (интернализованные Рц могут подвергаться дефосфорилированию и повторному встраиванию в плазматическую мембрану).
Введение опиоидного антагониста
Интернализация Рц, кроме того, может служить одним из путей деградации сигнального соединения.
У мышей с нокаутированным геном Р-аррестина-1 не наблюдается практически никаких отклонений от нормы. Нокаут гена р~ аррестина-2 приводит к выраженным изменениям реакции на агонист опиоидных Рц морфин: возрастают сила и длительность его действия и не развивается толерантность после повторных инъекций [в норме через 7 дней введения кривая «доза-ответ» (обезболивание) смещается приблизительно на порядок величины вправо]. Вместе с тем Р-аррестин-2 не участвует в развитии зависимости от морфина: введение опиоидного антагониста налоксона через 3 дня применения морфина вызывает симптомы отмены как у интактных мышей, так и у мышей с нокаутированным геном р-аррестина-2. Одновременный нокаут генов р-аррестина-1 и р-аррестина-2 приводит к гибели эмбрионов.
Независимые от G-белков эффекты GPCRs
Помимо отмеченной выше функции р-аррестинов в десенситизации и интернализации Рц, аррестины, по-видимому, могут выполнять функцию адапторных молекул, облегчающих активацию и внутриклеточную локализацию сигнальных каскадов (в частности, МАРК каскадов). Так, показано зависимое от агониста Рц взаимодействие р-аррестина с нерецепторной тирозинкиназой src28. Члены семейства src участвуют в регуляции пролиферации и дифференцировки клеток, активации каскада ERK и интернализации Рц за счёт фосфорилирования динамина и клатрина. Р-Аррестины, по-видимому, выполняют зависимую от агониста функцию «лесов», соединяющих последовательные киназы каскада МАРК: киназу киназы МАРК — МАРККК, киназу МАРК — МАРКК и МАРК (например, RAF1, МЕК1 и ERK или ASK1, МАРКК4 и JNK3). Белки-«леса» облегчают межбелковые взаимодействия, придают специфичность путям проведения сигнала и обеспечивают правильную внутриклеточную локализацию белков (например, перемещение МАРК из клеточного ядра в цитоплазму). Интересно, что р-аррестины прямо взаимодействуют с 1-м и 3-м членами каскада (т.е. МАРККК и МАРК), но не с промежуточным членом МАРКК.
Отсутствие агонистов
Оба р-аррестина выполняют такую функцию в отношении каскада ERK, но лишь р-аррестин-2 поддерживает каскад JUN3.
Ряд GPCRs содержит в С-концевой области мотив S/TxV(L/I), который узнаётся белками, содержащими домен PDZ. Например, Р2-АдРц взаимодействуют с содержащим PDZ регуляторным фактором обменника Na+—Н+, Рц серотонина 5-НТ2С — с белком MUPP1 (содержит множество доменов PDZ). С Р2-АдРц взаимодействует также содержащий БШ-домен адапторный белок Grb2, с Рц ангиотензина АТ1А — тирозинкиназа JAK2 через содержащую 8Н2-домен тирозинфосфатазу SHP. Рц могут также взаимодействовать с белками, содержащими SH3, WW или EVH домены, узнающие богатые пролином последовательности, а через них — с другими богатыми пролином белками, например, Рц Ins(l,4,5)P3 внутриклеточных депо Са2+. Независимо от G-белков, GPCRs могут взаимодействовать с малыми G-белками (например, Rho).
Роль GPCRs в патологии
Многие лиганды GPCRs стимулируют пролиферацию, а в ряде случаев (при избыточном образовании их Рц) и трансформацию клеток. Кроме того, обнаружен ряд активирующих мутаций Рц и G-белков, обеспечивающих активность Рц и нижележащих эффекторных систем и в отсутствие агонистов. Так, активирующие мутации Gas (блокирующие ГТФазную активность) могут вести к развитию токсичной аденомы и карциномы щитовидной железы, продуцирующей СТГ аденомы гипофиза, а мутации Ga12 — опухолей яичников и надпочечников. Активирующие мутации Рц ТТГ служат причиной развития гиперпродуцирующей тиреоидные гормоны аденомы щитовидной железы в 30% случаев этого заболевания. Активирующие мутации Рц ЛГ вызывают гиперплазию клеток Лейдига семенников и семейную форму преждевременного мужского полового созревания. Причиной опухолевого роста может также служить избыточная локальная продукция агонистов GPCRs (например, бомбезина, холецистокинина, вазопрессина и т.д.). Некоторые GPCRs кодируются геномом вирусов (например, цитомегаловируса человека).
- Предыдущие статьи из рубрики: Эндокринология
 Классификация ядерных рецепторов и их лиганды
Классификация ядерных рецепторов и их лигандыПредставители семейства ядерных Рц Представители семейства ядерных Рц могут предпочтительно взаимодействовать с тем или иным корепрессором. Помимо SMRT […]
 Общие свойства рецепторов
Общие свойства рецепторовРецепция и проведение сигнала Эти системы могут включать так называемые «вторые посредники» — низкомолекулярные соединения (циклические нуклеотиды, фосфолипиды […]
 Функции транспортных белков
Функции транспортных белковФермент печёночного типа Ферменты этой группы различаются между собой по субстратной предпочтительности в отношении эстрогенов и андрогенов и […]
 Транспорт сигнальных соединений
Транспорт сигнальных соединенийТранспортные белки и их лиганды К известным транспортным белкам относятся белки, связывающие стероиды (1), тиреоидные гормоны и ретиноиды […]
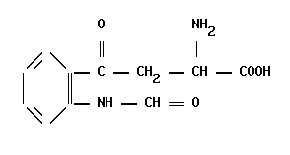 Производные триптофана
Производные триптофанаЗавершение процессов биосинтеза Так как конвертазы экспрессируются тканеспецифично, то и набор продуктов процессинга ПОМК в передней доле гипофиза, […]