
Функции транспортных белков
Фермент печёночного типа
Ферменты этой группы различаются между собой по субстратной предпочтительности в отношении эстрогенов и андрогенов и в отношении катализа в сторону окисления или восстановления. Мутации гена 17р-ГСД типа 3, экспрессия которого специфична для семенников, может вести к развитию мужского ложного гермафродитизма. 17р-ГСД типа 10 (в отличие от других ферментов этой группы локализованная на митохондриальной мембране) обладает также активностью L-3-гидроксиацил-КоА дегидрогеназы, т.е. может участвовать в обмене кетоновых тел и оказалась идентичной белку, связывающему р-пептид амилоида, играющего важную роль в патогенезе болезни Альцхаймера.
Для проявления биологической активности кортикостероидов важную роль играет 11р-гидроксил. Его окисление до кетогруппы ведёт к полной потере гормональной активности. Оксидоредукция в положении 11 катализируется двумя пр-гидроксистероид-дегидрогеназами. Фермент печёночного типа (тип 1) экспрессируется в чувствительных к глюкокортикоидам тканях и преимущественно катализирует восстановительную реакцию, т.е. способствует процессам реактивации гормонально неактивных 11-кетостероидов в активные кортизол и кортикостерон. Фермент почечного типа (типа 2), обнаруживаемый в чувствительных к альдостерону тканях, преимущественно катализирует обратную реакцию, т.е. способствует инактивации глюкокортикоидов. Данный изозим играет ключевую роль в дифференцированном действии глюко- и минералокортикоидов. Известно, что Рц минералокортикоидов одинаково хорошо взаимодействует с альдостероном и глюкокортикоидами. Учитывая, что концентрация глюкокортикоидов в крови на 2-3 порядка превышает концентрацию альдостерона, в отсутствие механизма дискриминации глюкокортикоидов альдостерон не мог бы выполнять своей специфической регуляторной функции. Функцию такого дискриминатора выполняет Іір-ГСД типа 2, инактивирующая глюкокор-тикоиды, но не влияющая на альдостерон, 11р-гидроксильная группа которого недоступна для фермента из-за образования 11—18 гемиацеталя.
Превращение андрогенов в эстрогены
 Превращение андрогенов в эстрогены под действием ароматазы в периферических тканях — минорный путь метаболизма андрогенов, но он служит важным источником эстрогенов в организме. Ароматаза экспрессируется во многих тканях, включая мозг, жировую ткань, мышцы. У грызунов перинатальная необратимая индукция ароматазы под действием андрогенов семенников в ряде областей мозга в значительной мере обеспечивает последующую половую дифференцировку нейроэндокринной системы и поведения. Образование эстрогенов в жировой ткани сглаживает переход к менопаузе у тучных женщин. Следует отметить, что 5а-дигидротестосте-рон является неароматизируемым андрогеном, и его применяют для выяснения эндокринного механизма действия тестостерона (прямого или через трансформацию в эстрогены).
Превращение андрогенов в эстрогены под действием ароматазы в периферических тканях — минорный путь метаболизма андрогенов, но он служит важным источником эстрогенов в организме. Ароматаза экспрессируется во многих тканях, включая мозг, жировую ткань, мышцы. У грызунов перинатальная необратимая индукция ароматазы под действием андрогенов семенников в ряде областей мозга в значительной мере обеспечивает последующую половую дифференцировку нейроэндокринной системы и поведения. Образование эстрогенов в жировой ткани сглаживает переход к менопаузе у тучных женщин. Следует отметить, что 5а-дигидротестосте-рон является неароматизируемым андрогеном, и его применяют для выяснения эндокринного механизма действия тестостерона (прямого или через трансформацию в эстрогены).
Наличие в периферических тканях Зр-ол-дегидрогеназы обеспечивает возможность активации секретируемого в значительных количествах надпочечниками дегидроэпиандростерона (DHEA) в цепочке превращений DHEA -> андростендион -» тестостерон -» 5а-дигидро-тестостерон.
Стероидные гормоны могут модифицироваться также путём дополнительного гидроксилирования под действием ряда цитохро-мов Р450. Данные превращения ведут к снижению или утрате гормональной активности стероидов, но, как правило, представляют минорный путь метаболизма. В то же время образование в гипоталамусе катехолэстрогенов при гидроксилировании эстрогенов в положениях 2 или 4 играет важную роль в системе положительной обратной связи в предовуляторный период. С одной стороны, кате-холэстрогены, обладая очень слабой эстрогенной активностью, но будучи эффективными лигандами Рц эстрогенов, блокируют систему отрицательной обратной связи эстрогенов на продукцию гонадолиберина. С другой стороны, катехолэстрогены эффективно конкурируют с индукторами секреции гонадолиберина катехолами-нами за фермент катаболизма катехоламинов — катехол-О-метил-трансферазу.
Эффективность стимулирующего действия
В результате эффективность стимулирующего действия катехоламинов на гонадолиберин возрастает. Катехолэстрогены через образование соответствующих хинонов способны образовывать аддукты ДНК, что может иметь отношение к известному проканцеявляется причиной синдрома CHILD (congenital hernidysplasia with ichthyosiform erythroderma and limb defects), летального для мужских особей. За-ГСД, относящиеся к семейству 1 альдо-кеторедуктаз, кодируемые минимально 4 генами, могут различаться по субстратной специфичности. Изозим С1 проявляет значительную 20а-ГСД активность (см. ниже). Изозим С2 обладает относительно низкой ферментативной активностью, но играет важную роль в транспорте жёлчных кислот в клетках печени. Изозим СЗ выполняет функцию синтазы PGF. В превращениях дигидропроизводных стероидов может также принимать участие 3-гидроксистероидэпимераза (семейство ретинолдегидрогеназ), способная катализировать эпимеризацию За-гидроксила через образование 3-кетогруппы. При этом из малоактивного 5а-андростан-За,17р-диола может образовываться активный андроген 5ос-дигидротестостерон, т.е. фермент способен осуществлять реактивацию андрогенов. Что касается кортикостероидов и прогестинов, то а- или р-восстановление 3-кетогруппы дополнительно снижает их гормональную активность. Однако тетрагидропроизвод-ные стероидов могут обладать и иными типами активности (например, активностью аллостерических регуляторов Рц-ионных каналов). У птиц 5р-дигидротестостерон — стимулятор гемопоэза.
Кортикостероиды и прогестины могут также подвергаться восстановлению по 20-кетогруппе под действием 20а- и 20р-гидро-ксистероиддегидрогеназ. Ряд гидроксистероиддегидрогеназ способен катализировать окислительно-восстановительные реакции по нескольким положениям стероидной молекулы. Например, один из ферментов печени кролика обладает За,3р,17р,20а-дегидрогеназными активностями. Ферментов, строго специфичных в отношении оксидоредукции стероидов в положении 20, по-видимому, нет. а- и р-Восстановление 20-кетогруппы кортикостероидов ведёт к резкому снижению гормональной активности.
Восстановленные прогестины
То же происходит при 20р-восстановлении прогестинов. В то же время 20а-восстановленные прогестины отчасти сохраняют гестагенную активность.
Характерной особенностью метаболизма андрогенов и эстрогенов является оксидоредукция кислородной функции в положении 17. В случае андрогенов окисление 17р-гидроксила до ке-тогруппы ведёт к полной утрате андрогенной активности, в то время как в группе эстрогенов такое превращение лишь ослабляет взаимодействие с Рц при сохранении полной агонистической гормональной активности. Реакции оксидоредукции катализируют 17р-гидро-ксистероидцегидрогеназы (ГСД). К настоящему времени клонировано не менее 10 таких ферментов. Ферменты этой группы различаются между собой по субстратной предпочтительности в отношении эстрогенов и андрогенов и в отношении катализа в сторону окисления или восстановления. Мутации гена 17Р-ГСД типа 3, экспрессия которого специфична для семенников, может вести к развитию мужского ложного гермафродитизма. 17Р-ГСД типа 10 (в отличие от других ферментов этой группы локализованная на митохондриальной мембране) обладает также активностью L-3-гидроксиацил-КоА дегидрогеназы, т.е. может участвовать в обмене кетоновых тел и оказалась идентичной белку, связывающему р-пептид амилоида, играющего важную роль в патогенезе болезни Алыгхаймера.
Для проявления биологической активности кортикостероидов важную роль играет Пр-гидроксил. Его окисление до кетогруппы ведёт к полной потере гормональной активности. Оксидоредукция в положении 11 катализируется двумя lip-гидроксистероид-дегидрогеназами.
- Предыдущие статьи из рубрики: Эндокринология
 Транспорт сигнальных соединений
Транспорт сигнальных соединенийТранспортные белки и их лиганды К известным транспортным белкам относятся белки, связывающие стероиды (1), тиреоидные гормоны и ретиноиды […]
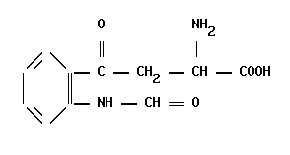 Производные триптофана
Производные триптофанаЗавершение процессов биосинтеза Так как конвертазы экспрессируются тканеспецифично, то и набор продуктов процессинга ПОМК в передней доле гипофиза, […]
 Тиреоидные гормоны
Тиреоидные гормоныЙодирование тироглобулина происходит в комплексе Гольджи у апикальной поверхности клеток под действием тиропероксидазы — гемсодержащего белка, катализирующего 1) […]
 Производные аминокислот
Производные аминокислотФосфолипазы А2 — большое семейство (известно около 20 членов) белков с различающейся субстратной предпочтительностью, локализацией (выделяют секретируемые, зависимые […]
- Биосинтез фосфолипидов
Биосинтез фосфолипидов группы РАР, обладающих сигнальными свойствами, может идти двумя путями. При биосинтезе de novo (преобладающем в почках […]